合肥肥料用Y氨基丁酸

- 面议
- 2025-01-25 04:08:05
- 肥料用Y氨基丁酸
- 兰海涛 13689608901
- 四川绵竹市鹏发生物科技有限责任公司

- 新浪微博


- QQ空间


- 豆瓣网


- 百度新首页

信息介绍
详细参数
在NO胁迫下的香蕉中也发现了GAD活性上升、GABA和香蕉多巴胺增加的现象。盐胁迫下谷氨酸脱氢酶活性与GAD的表达瞬时上升,进而提高GABA分流等相关途径的通量以调节碳氮平衡。应激下NADH:NAD+和 ADP:ATP的比值也能影响GABA-T,从而使GABA积累。盐胁迫下植物更多地利用C/N平衡途径缓解压力。
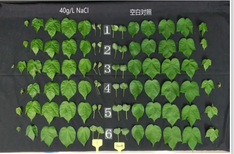
50mmol/L GABA和不同盐浓度会对植物幼苗产生不同的影响,当NO3-离子低于40mmol/L时,GABA会刺激 根伸长,当NO3-离子大于40mmol/L时GABA会抑制根伸长。并且GABA刺激低浓度的NO3-吸收,抑制高浓度NO3-的摄取,而GS等酶被氮调控,以上研究认为氮对调控植物生长有一定作用。在NaCl(50mmol/L)刺激下,植物的糖基化代谢会发起变化,并影响包括三羧酸循环、GABA代谢、氨基酸合成和莽草酸介导的次级代谢等发生变化。较高的盐离子会导致大豆的多胺氧化降解为GABA。植物GABA受体具有调节pH和Al3+的根耐受性。
细菌侵染过程中的植物GAD表达量和γ-羟基丁酸转录丰度会上升,致使GABA升高。高GABA合成水平的烟草对根癌土壤杆菌C58感染敏感性有所下降。GABA可诱导农杆菌ATTKLM操纵子表达,使得N-(3-氧代辛酰基)高丝氨酸内酯的浓度减少,群体感应信号(或激素)下调,影响其对植物的毒性。GABA在植物与细菌的信号交流中也发挥作用,GABA可以抑制细菌内Hrpl基因表达(Hrpl基因编码蛋白使得植物致敏或引起其组织疾病),同时抑制植物体内hrp基因表达,使得植物免于过敏反应(hrp:控制植物病原体致病能力,并引起过敏反应)。
GABA自动植物以及微生物中有较多的发现,其中在1949年在马铃薯的块茎中发现,在1950年又在哺乳动物的系统中发现其存在,同时被认为是哺乳动物、昆虫或者某些寄生蠕虫神经系统中的神经抑制剂,对神经元的兴奋程度有着重要的影响。
植物富集法是一种新型开发的合成萃取提纯技术,它是用GABA含量较高的植物进行分离提取,这样便有了既便宜纯度又高的GABA产品。从植物中获取GABA的方法主要有以下两种:其中一种是利用溶剂萃取提纯法,另一种是柱分离制备法。溶剂萃取法,溶剂萃取法是利用水或醇作为GABA的提取剂,根据植物在水或者醇中的溶解度以及分配系数不同的原理将GABA提出到水或者醇中,并且经过反复的过滤提纯,可以使植物中绝大多数的GABA都被萃取出来。柱分离制备法,柱分离制备法,又被叫做柱色谱法,是一种利用不同的混合物中的组分在固液两相中具有不同分配系数的原理,进行洗脱分离及其他后续操作,它的大分类应该归属于层析法。色谱柱一般采用树脂、硅胶或活性炭等作为填充材料。
在高等植物中,GABA的代谢主要由三种酶参与完成,在GAD作用下,L-谷氨酸(glutamic acid,Glu)在α-位上发生不可逆脱羧反应生成GABA,然后在GABA转氨酶(GABA transaminase,GABA-T)催化下,GABA与丙酮酸和α-酮戊二酸反应生成琥珀酸半醛,后经琥珀酸半醛脱氢酶(succinic semialdehyde dehydrogenase,SSADH)催化,琥珀酸半醛氧化脱氢形成琥珀酸终进入三羧酸循环(krebs circle)。这条代谢途径构成了TCA循环的一条支路,称为GABA支路。
- 鹏发
- 肥料用Y氨基丁酸
- 氨基酸类衍生物添加剂
- 兰海涛


